Содержание
Аномальная активность
Гипотония может быть вызвана повреждением альфа-нейронов или афферентов Ia, несущих сенсорную информацию альфа-нейронам. Это создает снижение мышечного тонуса. Напротив, гипертония вызывается повреждением нисходящих путей, которые заканчиваются в спинном мозге. Он увеличивает мышечный тонус за счет увеличения общей реакции альфа-мотонейронов на сенсорный вход Ia.
Спазмы могут быть вызваны несоответствием между активностью альфа- и гамма-мотонейронов, т. Е. Слишком большим усилением одного или другого. Дисбаланс вызывает неточные показания мышечных рецепторов в мышечном веретене. Следовательно, сенсорные нейроны, ответственные за головной и спинной мозг, вводят в заблуждение. Например, если у пациента слишком активны гамма-двигательные нейроны, будет сопротивление пассивному движению, вызывающее скованность, также называемую спастичностью . Это часто встречается у людей с повреждением высших центров, влияющих на нисходящие пути. Иногда это может привести к тому, что гамма-смещение (постоянный разряд некоторых гамма-мотонейронов) будет больше или меньше обычного. В случае пациентов с избыточным гамма-смещением сенсорные окончания в мышечных веретенах слишком часто разряжаются, что приводит к большей мышечной активности, чем необходимо. Кроме того, эта гиперактивность в гамма-петле веретена может вызвать спастичность.
Гамма-мотонейроны помогают удерживать мышечное веретено в напряжении, тем самым регулируя чувствительность. Следовательно, если не происходит надлежащего возбуждения гамма-мотонейронов, движение мышц может быть нарушено. Больше всего страдают мелкие моторные навыки, такие как движения пальцами и глазами, поскольку отсутствие напряжения в мышечном веретене препятствует его способности определять степень растяжения через сенсорные окончания. Это означает, что мышца не сможет точно двигаться соответственно. Поражения, контролирующие нисходящие пути от нижних мотонейронов к верхним конечностям, могут привести к потере способности пациента точно контролировать движения.
В клинических условиях можно проверить, имеет ли кто-то аномально низкое или высокое усиление гамма-излучения, просто перемещая руку пациента. Гамма-усиление — это процесс, при котором ускорение, скорость и длина мышечных изменений увеличиваются в равной степени, что позволяет выполнять более точные движения в соответствующей ситуации. Если сложнее согнуть руку пациента в локте вперед и назад, тогда он / она имеет большее усиление гаммы, в то время как тот, чья рука очень легко двигается, будет иметь меньшее усиление гаммы.
Осциллографы могут использоваться для измерения потенциалов действия аксона от мотонейрона с целью оценки общей мышечной активности. Хотя он не может отличить альфа-мотонейроны от гамма-мотонейронов, он полезен для понимания того, есть ли у одного из них аномальная активность мотонейронов. При низкой скорости активности нисходящего пути активируется меньшее количество мотонейронов меньшего размера, что приводит к небольшой мышечной силе. Это будет отображаться на осциллографе в виде более низких пиков на оси ординат.
Медикаментозная терапия
Этиотропного лечения мотонейронных поражений не существует. Надежды в достижении клинического эффекта у пациентов связывают с патогенетической терапией. Целью лечения является замедление развития патологического процесса и уменьшения выраженности его симптомов
Важно начать лечение раньше
Эффект наблюдается при применении пресинаптического ингибитора высвобождения медиатора возбуждения глутамата, препарат рилутек. В качестве метода лечения назначают комплекс витаминов группы В. Их применение даёт возможность воздействовать на комплекс формирующихся вегетативных, сенсорных и трофических нарушений.
Связи[править | править код]
Как и другие нейроны, нижние двигательные нейроны обладают как афферентными (входящими) и эфферентными (исходящими) связями. Альфа-мотонейроны получают афферентную иннервацию из ряда источников, в том числе верхних мотонейронов, сенсорных нейронов и интернейронов. В свою очередь α-МН иннервируют экстрафузальные мышечные волокна. Эти афферентные и эфферентные связи участвуют в координации мышечной активности.
Афферентный входправить | править код
| Расположение ВМН | Расположение α-МН | Имя пути |
|---|---|---|
| Головной мозг | Ствол мозга | Корково-ядерный путь |
| Головной мозг | Спинной мозг | Пирамидный тракт |
| Красное ядро | Спинной мозг | Красноядерно-спинномозговой тракт |
| Вестибулярные ядра | Спинной мозг | Преддверно-спинномозговой тракт |
| Крыша среднего мозга | Спинной мозг | Покрышечно-спинномозговой тракт |
| Ретикулярная формация | Спинной мозг | Ретикулярно-спинномозговой тракт |
Верхние мотонейроны (ВМН) посылают аксоны к α-МН через несколько путей, включая (но не ограничиваясь) корково-ядерный, пирамидный и красноядерно-спинномозговой проводящие пути.
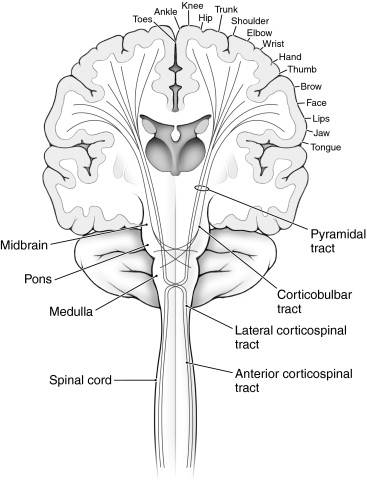 Анатомия проводящих путей нервной системы
Анатомия проводящих путей нервной системыКорково-ядерный путь соединяет кору больших полушарий с ядрам черепных нервов. (Корково-ядерный путь также называют кортикобульбарным путём.) Он представляет собой пучок аксонов верхних двигательных нейронов, который спускается из коры и оканчивается синапсами на α-МН ствола мозга. Аналогично, ВМН коры головного мозга напрямую контролируют α-МН в спинном мозге через латеральные и вентральные корково-спинномозговые тракты.
Афферентный (сенсорный) вход для α-МН обширен и берет своё начало в сухожильных органах Гольджи, мышечных веретенах, механорецепторах, терморецепторах, и других сенсорных нейронах на периферии. Эти связи образуют структуру нейронных цепей, которые лежат в основе рефлексов. Существует несколько типов рефлекторных схем, простейшая из которых состоит из одного синапса между сенсорным нейроном и α-МН. Коленный рефлекс является примером такого моносинаптического рефлекса.
Наиболее значительная афферентация приходит на α-МН от локальных интернейронов, которые являются наиболее многочисленным типом нейронов в спинном мозге. Помимо множества других функций которые они выполняют, интернейроны посылают аксоны к α-МН, что создаёт более сложные рефлекторные схемы. Одним из типов интернейронов являются клетки Реншоу, которые будут рассмотрены ниже.
Эфферентный выходправить | править код
Альфа-мотонейроны посылают аксоны, которые в основном оканчиваются синапсами на экстрафузальных мышечных волокнах. Некоторые аксоны α-МН оканчиваются на клетках Реншоу, т. е. тормозных интернейронах, которые посылают аксоны к α-МН и ограничивают их активность с целью предотвращения повреждения мышц.
Диагностика и лечение
При первых проявлениях патологии нужно немедленно обратиться к специалисту и пройти полное обследование. Если пациент отмечает у себя прогрессирующую генерализованную двигательную слабость, которая параллельно сопровождается значительными отклонениями чувствительности, то это первые тревожные признаки заболевания.
Обследование включает в себя проведение:
- Игольчатая ЭМГ. Считается информативным методов.
- Исследование скорости проведения возбуждения.
- Лабораторные тесты. Обязательно сдается общий анализ крови, чтобы определить уровень электролитов, белка, а также гормонов щитовидной железы.
- Магнитно-резонансная томография шейного отдела позвоночника. Проводится только в том случае, если отсутствуют клинические и ЭМГ данные, которые свидетельствуют о поражении черепных нервов.
Также пациенту может назначаться и люмбальная пункция. Параллельно сдается анализ на наличие венерических инфекций, определяют уровень СОЭ. Если пациент заинтересован в генетическом консультировании, то проводится и генетическое тестирование.
Что касается лечения, то специфической терапии не существуют. Пациенту прописывают:
- «Рилузол», который необходимо принимать регулярно по два раза в сутки. Помогает облегчить бульбарную форму бокового амиотрофического склероза.
- «Баклофен».
- «Гликопирропат», «Амтриптилин».
- «Флувоксамин».
На более поздних стадиях патологии могут прописать опиоиды и бензодиазепины.
 Спинальные рефлексы: классификация, функции и свойства
Спинальные рефлексы: классификация, функции и свойстваЧтобы помочь пациенту побороть неврологические отклонения данного характера, стоит обратить внимание также на такие методы:
- Физиотерапия. Поддерживает мышечную функцию. Рекомендуется использовать ортопедические бандажи, которые обладают фиксирующей функцией.
- Консультация и занятия с логопедом. Помогает подобрать коммуникативные устройства, которые облегчат процесс общения.
- Чрескожная эндоскопическая гастростомия.
- Неинвазивная дыхательная поддержка. Назначается при респираторной слабости.
Хирургическое вмешательство для облегчения глотания проводится в редких случаях, так как считается малоэффектинвым.
Болезнь мотонейрона – серьезная неврологическая патология, которая приводит не только к серьезным последствиям, но даже к летальному исходу. Желательно проводить своевременную диагностику и лечение.
Перспективы лечения
Подход к коррекции палогического состояния должен быть комплексным. Сюда относится изменение режима и качества питания, кардио тренировки, регулярные физические нагрузки и лекарственная терапия. Важную роль играет и психологическая коррекция, которая направлена на формирование механизмов адаптации и является обязательной составной частью ведения больных с болезнью мотонейронов. При депрессивных явлениях рекомендуют консультацию психолога с дальнейшим назначением начальной дозы транквилизаторов.
Коррекция режима питания обусловлена тем, что при патологии нарушается акт глотания. Диетолог поможет правильно подобрать меню. При фарингеальной слабости прием пищи представляет затруднение, поэтому может понадобиться эндоскопическая гастростомия.
Необходима будет и консультация инструктора по лечебной физкультуре. Для разработки мышц, поддержании их в тонусе пациентам рекомендуют постоянные физические упражнения, плавание, занятие индивидуально с тренером в спортивном зале.
Развитие
Гамма-моторные нейроны происходят из базальной пластинки эмбриона.
Гамма-мотонейроны вначале развиваются аналогично альфа-мотонейронам. Они берут начало в базальной пластинке , которая является вентральной частью нервной трубки развивающегося эмбриона . Гены Sonic hedgehog (Shh) — важная часть процесса развития, секретируемая хордой, создающей градиенты концентраций. После генов hedgehog, различные другие молекулярные маркеры и факторы транскрипции играют роль в дифференциации мотонейронов в специфические гамма-мотонейроны.
 Возрастные особенности нервной системы человека
Возрастные особенности нервной системы человекаГамма-мотонейроны, как и все клетки, при рождении экспрессируют определенные генетические маркеры. Нейротрофические факторы GDNF, полученные из мышечного веретена, также должны присутствовать для постнатального выживания. Wnt7A представляет собой секретируемую сигнальную молекулу, избирательно секретируемую в гамма-мотонейронах на 17,5-й день эмбриона мышей. Это самая ранняя молекула, присутствующая в гамма-мотонейронах, которая отличает их от альфа-мотонейронов, иллюстрируя дивергенцию этих двух типов нижних мотонейронов.
Кроме того, было сделано заключение, что рецептор серотонина 1d (5-ht 1d) является новым маркером гамма-мотонейронов, позволяющим исследователям различать различные типы нижних мотонейронов . Мыши, лишенные этого рецептора серотонина 1d, демонстрировали более низкий моносинаптический рефлекс ( рефлекторная дуга, включающая только сенсорный и двигательный нейрон), что может быть вызвано сниженным ответом на сенсорную стимуляцию моторных нейронов. Кроме того, мыши с нокаутом без этого рецептора серотонина проявляли большую координацию при выполнении задачи с балансиром, предполагая, что меньшая активация моторных нейронов афферентами Ia во время движения может уменьшить ненужный избыток мышечной продукции.
Другой отличительный молекулярный маркер гамма-мотонейронов — фактор транскрипции Err3. Он экспрессируется на высоком уровне в гамма-мотонейронах, но очень мало в альфа-мотонейронах. С другой стороны, нейрональный ДНК-связывающий белок NeuN присутствует в значительно больших количествах в альфа-мотонейронах. Остеопонтин , белок, также экспрессирующийся в костях, отсюда и префикс «остео-», является маркером альфа-мотонейронов. Это, в свою очередь, может предоставить ученым способ устранить гамма-мотонейроны, если интересуют альфа-мотонейроны. Одно исследование, в частности, сделало этот вывод, основываясь на том факте, что остеопонтин присутствует в более крупных клеточных телах, что указывает на альфа-мотонейроны, поскольку они имеют более крупные клеточные тела, чем гамма-мотонейроны.
Мышечный тонус
Хотя мускулы могут находиться в расслабленном состоянии, у них есть общий уровень напряжения покоя. Это называется мышечным тонусом и поддерживается двигательными нейронами, иннервирующими мышцу. Его цель — поддерживать осанку и способствовать более быстрым движениям, поскольку, если бы мышцы были полностью расслаблены, потребовалось бы больше нервных импульсов.
Величина напряжения в мышцах зависит в первую очередь от разряда альфа-мотонейронов на уровне покоя, в основном афферентов веретена Ia. Гамма-мотонейроны также задействованы посредством своего действия на интрафузальные мышечные волокна. Интрафузальные мышечные волокна контролируют уровень покоя афферентного пути Ia, что, в свою очередь, создает устойчивый уровень активности альфа-нейронов.
Мышечный тонус также может быть обусловлен тоническими разрядами гамма-мотонейронов. Активация этих нейронов происходит в основном из нисходящих волокон фасилитирующей ретикулярной формации . Это приводит к растяжению мышечного веретена, активации альфа-мотонейронов и, наконец, к частично сокращенной мышце. Мозжечок является связующим звеном альфа-гамма двигательных нейронов. Следовательно, в мозжечке мышечное напряжение поддерживается с помощью альфа-мотонейронов, а также гамма-мотонейронов.
Клиника основных форм заболевания
Характерно 4 формы:
- боковой амиотрофический склероз;
- прогрессирующий бульбарный паралич;
- мышечная атрофия с прогрессирующим течением;
- первичный боковой.
Клиника зависит от локализации поражения и степени выраженности дегенеративных изменений.
Проявления бокового амиотрофического склероза
Признаки поражения нижнего мотонейрона:
- мышечная слабость;
- снижение рефлексов с конечностей;
- атрофия мышц;
- фасцикуляции (подёргивания);
Первая жалоба таких больных — слабость мышц кисти и болезненные спазмы при потягивании в момент пробуждения. Затем появляется затруднение акта жевания и глотания. При поражении диафрагмы больной может погибнуть еще до развития полной клинической картины болезни.
 БАС является прогрессирующим заболеванием центральной нервной системы с обязательным поражением диафрагмы, что приводит к нарушению функции внешнего дыхания и полной его остановке.
БАС является прогрессирующим заболеванием центральной нервной системы с обязательным поражением диафрагмы, что приводит к нарушению функции внешнего дыхания и полной его остановке.
Скорость прогрессирования болезни может быть быстрой с развитием дыхательной недостаточности в течение года, средней (от 2-х до пяти лет) и медленной. В далеко зашедшей стадии может наблюдаться задержка или недержание мочи.
Прогрессирующий бульбарный паралич
Болезнь распространяется на нейроны спинного и головного мозга, проявляется нарастающими затруднениями в жевании, глотании, неразборчивости речи. Разговор таких пациентов становится нечётким, появляется гнусавость и охриплость голоса.
Обращает на себя внимание отсутствие мимики на лице. Во время приёма пищи еда может выпадать из полости рта
При парезе дыхательной мускулатуры развивается летальный исход.
Прогрессирующая мышечная атрофия
Является наследственным заболеванием. Первые симптомы развиваются в любом возрасте и проявляются мышечными подергиваниями, локальными судорогами и фасцикуляциями – спонтанными и одновременными сокращениями одной мышечной группы.
Первичный латеральный склероз
Клиническая картина этой нозологии характеризуется повышением тонуса мышц нижних конечностей, нарушением ходьбы и координации.
Таким пациентам трудно сохранять равновесие. У больных изменяется голос, характер речи и глотание, а к концу заболевания возникают трудности при дыхании.
Причины гибели нейронов
Заболевание мультифакториальное, т.е. его развитие происходит из-за совокупности влияния наследственных факторов и окружающей среды.
Основные факторы развития болезни мотонейрона:
- Мутация генов, расположенных в 21 паре хромосом.
- Наличие четырёх спиральных ДНК и РНК.
- Активный аутоиммунный процесс.
- Болезни паращитовидных желез.
- Избыточное содержание медиаторов возбуждения.
- Перенесенная вирусная инфекция, оказывающая токсическое воздействие на двигательные нейроны.
- Избыточное содержание ионов кальция в клетках и накопление большого количества свободных радикалов.
- Влияние экзотоксинов, в том числе никотин.
- Наличие сопутствующих заболеваний (гипертоническая болезнь, сахарный диабет второго типа, аденома простаты, доброкачественные опухоли).
Связь
Как и другие нейроны, нижние двигательные нейроны имеют как афферентные (входящие), так и эфферентные (исходящие) связи. Альфа-мотонейроны получают входные данные от ряда источников, включая верхние мотонейроны , сенсорные нейроны и интернейроны . Первичный выход α-МН — экстрафузионные мышечные волокна . Эта афферентная и эфферентная связь необходима для достижения скоординированной мышечной активности.
Афферентный ввод
| UMN происхождение | α-MN мишень | Название тракта |
|---|---|---|
| Кора головного мозга | Мозговой ствол | Кортикоядерный тракт |
| Кора головного мозга | Спинной мозг | Кортикоспинальный тракт |
| Красное ядро | Спинной мозг | Руброспинальный тракт |
| Вестибулярные ядра | Спинной мозг | Вестибулоспинальный тракт |
| Тектум среднего мозга | Спинной мозг | Тектоспинальный тракт |
| Ретикулярная формация | Спинной мозг | Ретикулоспинальный тракт |
Верхние мотонейроны (UMN) посылают входные данные в α-MN несколькими путями, включая (но не ограничиваясь ими) кортиконуклеарный , кортикоспинальный и руброспинальный тракты . Кортикоядерный и кортикоспинальный тракты обычно встречаются при исследованиях взаимодействия верхних и нижних мотонейронов при контроле произвольных движений.
Кортико — нуклеарный тракт назван так потому , что она соединяет кору головного мозга к черепным ядрам нервных . (Кортиконуклеарный тракт также называется кортикобульбарным трактом , поскольку мишень в стволе мозга — продолговатом мозге — архаично называется «луковицей».) Именно по этому пути верхние мотонейроны спускаются из коры и синапсов на α-МН. ствола мозга. Точно так же UMN коры головного мозга напрямую контролируют α-MN спинного мозга через латеральные и вентральные кортикоспинальные тракты .
Сенсорный ввод в α-МН обширен и берет свое начало в органах сухожилий Гольджи , мышечных веретенах , механорецепторах , терморецепторах и других сенсорных нейронах на периферии. Эти связи обеспечивают структуру нейронных цепей, лежащих в основе рефлексов . Существует несколько типов рефлекторных цепей, самый простой из которых состоит из одного синапса между сенсорным нейроном и α-МН. Коленный рефлекс является примером такого моносинаптического рефлекса.
Самый обширный вход в α-МН поступает от локальных интернейронов , которые являются наиболее многочисленным типом нейронов спинного мозга . Среди множества их функций — синапсы интернейронов на α-MN для создания более сложных рефлекторных схем. Одним из типов интернейронов является клетка Реншоу .
Эфферентный выход
Альфа-мотонейроны посылают волокна, которые в основном синапсы, на экстрафузные мышечные волокна . Другие волокна от синапса α-MN на клетках Реншоу , то есть тормозящие интернейроны, которые синапсируют с α-MN и ограничивают его активность, чтобы предотвратить повреждение мышц.
Развитие
Моторные нейроны начинают развиваться на ранних этапах эмбрионального развития , и двигательные функции продолжают развиваться в детстве. В клетки расположены либо на рострально-каудальной оси, либо на вентрально-дорсальной оси. В аксоны двигательных нейронов начинают появляться на четвертой неделе развития из вентральной области вентральной-спинные оси ( донца ). Этот гомеодомен известен как домен предшественника моторных нейронов (pMN). Факторы транскрипции здесь включают Pax6 , OLIG2 , Nkx-6.1 и Nkx-6.2 , которые регулируются sonic hedgehog (Shh). Ген OLIG2 является наиболее важным из-за его роли в стимулировании , гена, который вызывает выход из клеточного цикла, а также способствует дальнейшим факторам транскрипции, связанным с развитием моторных нейронов.
Дальнейшая спецификация мотонейронов происходит, когда ретиноевая кислота , фактор роста фибробластов , Wnts и TGFb интегрируются в различные факторы транскрипции Hox . Существует 13 факторов транскрипции Hox, которые вместе с сигналами определяют, будет ли мотонейрон более ростральным или каудальным по характеру. В позвоночнике Hox 4-11 сортируют мотонейроны в один из пяти моторных столбцов.
| Моторная колонка | Расположение в спинном мозге | Цель |
| Средняя колонка двигателя | Присутствует вся длина | Осевые мышцы |
| Гипаксиальная моторная колонка | Грудной отдел | Мышцы стенки тела |
| Преганглионарная моторная колонка | Грудной отдел | Симпатический ганглий |
| Боковая моторная колонка | Плечевая и поясничная области (обе области дополнительно делятся на медиальную и латеральную области) | Мышцы конечностей |
| Диафрагмальная моторная колонка | Шейный отдел | Диафрагма |
Место расположения
Альфа-мотонейроны (α-МН), иннервирующие голову и шею, находятся в стволе мозга ; остальные α-МН иннервируют остальную часть тела и находятся в спинном мозге . В спинном мозге больше α-МН, чем в стволе мозга, поскольку количество α-МН прямо пропорционально степени контроля мелкой моторики в этой мышце. Например, мышцы одного пальца имеют больше α-MN на волокно и больше α-MN в целом, чем мышцы четырехглавой мышцы , что позволяет более точно контролировать силу, прилагаемую пальцем.
Как правило, альфа-МН на одной стороне ствола или спинного мозга иннервируют мышцы на той же стороне тела. Исключением является блокирующее ядро в стволе мозга, которое иннервирует верхнюю косую мышцу глаза на противоположной стороне лица.
Мозговой ствол
В стволе мозга α-МН и другие нейроны располагаются в кластерах клеток, называемых ядрами , некоторые из которых содержат тела нейронов, принадлежащих черепным нервам . Не все ядра черепных нервов содержат α-МН; те, что есть, являются двигательными ядрами , а другие — сенсорными ядрами . Ядра двигателя находятся по всему brainstem- мозговом , моста и среднего мозга й по причинам развития находятся вблизи средняя линии мозга.
Как правило, двигательные ядра, расположенные выше в стволе мозга (то есть более ростральные), иннервируют мышцы, расположенные выше на лице. Например, глазодвигательное ядро содержит α-МН, которые иннервируют мышцы глаза, и находится в среднем мозге, самом ростральном компоненте ствола мозга. Напротив, подъязычное ядро , которое содержит α-МН, иннервирующие язык, находится в продолговатом мозге, наиболее каудальной (т. Е. В нижней части) структур ствола мозга.
Спинной мозг
Кортикоспинальный тракт является одним из основных нисходящих путей от мозга к альфа-МНБУ спинного мозга .
В спинном мозге α-МН расположены в сером веществе , образующем вентральный рог . Эти α-МН обеспечивают двигательный компонент спинномозговых нервов, которые иннервируют мышцы тела.
Альфа-мотонейроны расположены в ламине IX согласно системе пластинок Рекседа .
Как и в стволе головного мозга, более высокие сегменты спинного мозга содержат α-МН, которые иннервируют мышцы выше на теле. Например, двуглавая мышца плеча , мышца руки, иннервируется α-МН в сегментах C5, C6 и C7 спинного мозга, которые находятся рострально в спинном мозге. С другой стороны, икроножная мышца , одна из мышц ноги, иннервируется α-МН в сегментах S1 и S2, которые находятся в спинном мозге каудально.
Альфа-мотонейроны расположены в определенной области серого вещества спинного мозга. Эта область обозначается lamina IX в системе пластинок Rexed , которая классифицирует области серого вещества на основе их цитоархитектуры . Lamina IX располагается преимущественно в медиальной части вентрального рога, хотя есть некоторый вклад в lamina IX за счет набора двигательных нейронов, расположенных более латерально. Как и в других областях спинного мозга, клетки в этой пластинке соматотопически организованы, а это означает, что положение нейронов в спинном мозге связано с тем, какие мышцы они иннервируют. В частности, α-МН в медиальной зоне ламины IX имеют тенденцию иннервировать проксимальные мышцы тела, тогда как те, что в боковой зоне, имеют тенденцию иннервировать более дистальные мышцы. Аналогичная соматотопия связана с α-MNs, которые иннервируют мышцы-сгибатели и разгибатели: α-MNs, которые иннервируют сгибатели, как правило, расположены в дорсальной части lamina IX; те, которые иннервируют разгибатели, обычно расположены более вентрально.

 Физиология спинного мозга, ретикулярной формации, спинальный шок
Физиология спинного мозга, ретикулярной формации, спинальный шок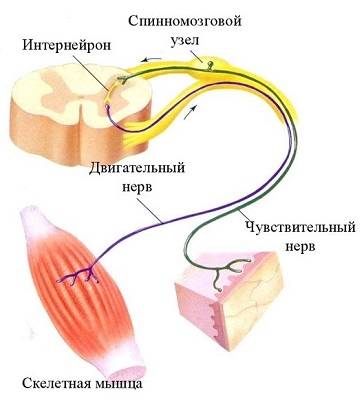 Рефлекторная дуга в биологии
Рефлекторная дуга в биологии Проводящие пути головного и спинного мозга
Проводящие пути головного и спинного мозга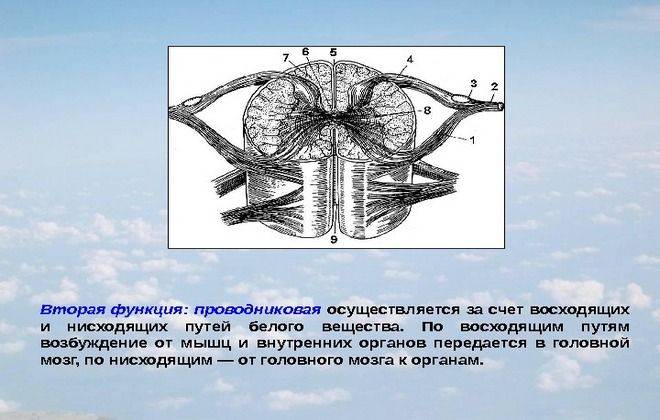 Спинной мозг строение и функции кратко
Спинной мозг строение и функции кратко